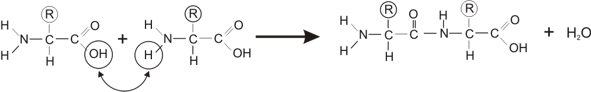
But: étudier les lois qui régissent le fonctionnement de la cellule.
Outils: Les biotechnologies
Historique:
➢ Deuxième moitié du 20
ème
siècle: invention du microscope.
➢ Hooke en 1650 observe des coupes de Liège et observe une cellule
➢ Schleiden et Schwann en 1830 établissent les 2 premières règles de la biologie cellulaire:
• Tous les organismes sont constitués de plusieurs cellules.
• La cellule est l'unité fondamentale de la vie.
➢ Virchow établit 30 ans plus tard la 3ième règle de la biologie cellulaire:
• Une cellule est créée à partir d'une cellule préexistante.
➢ Zeist: fin XIX
ème
siècle fait évoluer le microscope optique (MO) avec la lentille.
➢ 1930: Microscope interférenciel permettant d'observer des cellules vivantes
➢ 1930: Microscope électronique (ME)
➢ 1945: Première description des organites cellulaires
➢ 1945: Immunofluorescence
➢ 1988: Microscope confocal
Le résultat final dépend de l’outil et de l’interprétation faite à partir de l’observation
Il existe deux types de cellules, différenciées par la présence ou non de matériel génétique dans un
noyau:
Les Eucaryotes ont le matériel génétique dans un noyaux, les cellules eucaryotes sont unis
et pluri-cellulaires
Les Procaryotes dont le matériel génétique est libre. Ce sont des organismes unicellulaires.
Structure générale d’une cellule eucaryote:
PCEM1 - Biologie Cellulaire 1/10 www.ProteinesPlus.frCours 01 - Les méthodes d'observation en Biologie Cellulaire
La membrane cytoplasmique joue plusieurs rôles:
• Barrière vis-a-vis de l'environnement mais perméable: il y a passage selectif de divers
ions
• Permet l'absorption d'élément extracellulaire par invagination ou endocytose
• Permet l'excretion de substance intra-cellulaire par exocytose.
• Zone d'interaction avec l'environnement qui se fait de différentes manières:
• la membrane présente des récepteurs où peuvent se fixer des liguands qui
déclencheront des réactions
• la zone d'adhérence à la MeC ou a une autre cellule voisine
• Forme des expansions grâce au cytosquelette aux rôles variés: déplacement et
detection de l'environnement.
Dans le noyau: l'ADN est sous forme de chromatine dense (hétérochromatine) ou
chromatine claire (euchromatine).
Il présente une condensation à contenu importante en ARN: le nucléole.
Le noyau a une double membrane perforée permettant une communication entre le
nucléoplasme et le cytoplasme. Les échanges se feront dans les deux sens
Dans le noyau se fait la transcription en ARN qui passe dans le cytoplasme où il sera traduit
en Protéines grâce aux Ribosomes.
Le fond cytoplasmique est constitué de Cytosol ou Hyaloplasme, qui est un gel dans
lequel sont dissous un certains nombres de molécules et ou vont se trouver les differents
organites cellulaires.
Les éléments membranaires:
• Réticulum endoplasmique granuleux (REG) présentant des ribosomes à sa
surface, est responsable de la synthèse des protéines membranaires, les protéines des
lysosomes ainsi que les protéines destinées à l'exportation
Les Ribosomes libres synthètisent des protéines qui resteront dans le cytoplasme. Ce
qui est synthétisé dans le REG sera empacté dans l'appareil de Golgi.
• L’appareil de Golgi est un empilement de saccules formant les Lysosomes (appareil
de digestion) ou grain de secrétion dont le contenu est destiné à l'exportation
• Le système endosomique: Ensemble du système qui forme le carrefour entre ce qui
arrive de la Mb cytoplasmique et ce qui dérive de Golgi. On parle de système endolysosomal.
• Les mitochondries sont à l'origine de la respiration cellulaire et de la production
d'énergie, son système membranaire comporte des Peroxysomes.
Le cytosquelette est l'ensemble de filaments et tubes formés par des protéines qui
s'associent entres elles. Il est composé de: Microfilaments d'actine, Filaments
Intermédiaire, Microtubules.
Cette architecture cellulaire est fortement lié au fonctionnement intra-cellulaire:
ex: une cellule qui produit beaucoup de protéines aura beaucoup de ribosomes.
Ces structures sont plus ou moins développées selon la fonction de la cellule observée.
corrélation entre la morphologie cellulaire et la fonction de la cellule.
Morphologie/Structure <=> Fonction/Metabolisme <=>Environnement
PCEM1 - Biologie Cellulaire 2/10 www.ProteinesPlus.frCours 01 - Les méthodes d'observation en Biologie Cellulaire
I. Les Microscopes Optiques (MO)
1. Microscope optique (MO) ou photonique
Il est constitué de bas en haut:
• La lumière placée tout en bas.
• Le condenseur qui condense la lumière sur la préparation
• La platine sur laquelle repose l'échantillon
• L'objectif qui permet d'agrandir l'image 4-10-40x
• L'occulaire qui agrandit 10 x l'image.
Le grossissement va donc de 40 à 400x. Les contraintes sont établies par:
• La qualité des lentilles de verre
• La capacité de la lumière à traverser l'échantillon à observer.
La préparation doit donc être fine:
Dans le cas d'un étalement cellulaire (frotti sanguin): on dépose la goutte de sang sur une
lame, et à l'aide d'une lamelle on étale la goutte jusqu'à obternir une monocouche de 1-2
microns d'épaisseur.
Pour une culture cellulaire: les cellules se multiplient et le cytoplasme est assez fin.
En cas de prélèvement de tissus, celui ci doit être coupé une fois durci, et cela peut se faire
de deux manière:
• Congélation du tissu qui pourra ensuite être coupé à l'aide d'un Cryostat.
• Enrobage dans une inclusion de Paraffine, ce qui est plus long et peut détruire les
cellules si on ne les fixe pas au Formaldéhyde ou Formol qui va stabiliser les
protéines en formant des liens entre elles. Les Lipides eux ne seront pas conservés.
La paraffine est solide à température ambiante, mais est liquide à 55°C, insoluble dans l'eau. Ainsi,
le prélèvement doit être d'abord baigné dans des solutions d'éthanol à concentrations de plus en
plus élevés. L'alcool se substituera à l'eau.
Cependant, la paraffine n'est pas totalement miscible avec l'éthanol. L'échantillon sera alors baigné
dans du Toluène, après quoi il sera baigné dans de la paraffine qui va elle même se substituer au
toluène.
Le bloc sera coupé à l'aide d'un Microtome qui est un appareil constitué d'un support sur lequel
repose l'échantillon sur lequel passe la lame d'un couteau.
L'épaisseur de la coupe est d'environ 4-8 microns. Ces coupes seront transferées sur une lame.
Cependant, cette coupe est tellement fine qu'il y a trop peu de contraste, il faudra alors la
colorer à l'hématoxyline-éosine (hématoxyline pour le noyau, éosine pour le cytoplasme et la MeC)
par exemple. Mais on ne peut pas colorer la lame puisqu'enrobée par la paraffine.
On suit alors le procedé inverse: déparaffination et réhydratation.
On va alors proteger cette coupe en la mettant dans un milieu de montage qui doit être
transparent, et qui sera soit acqueux soit non miscible à l'eau mais avec les solvants (et
donc refaire la déshydratation).
Ces cellules sont figées/mortes: aucune activité ne pourra être relevée.
2. Microscope à contraste de phase
Il utilise le fait que lorsque la lumière traverse la cellule, elle rencontre des structures qui
vont devier ses ondes, donc celles-ci vont sortir soit en phase, soit déphasées.
Cette différence sera à l'origine d'un léger contraste qui fera apparaître le noyau et les
chromosomes.
Elle permet l'étude de cellules vivantes.
PCEM1 - Biologie Cellulaire 3/10 www.ProteinesPlus.frCours 01 - Les méthodes d'observation en Biologie Cellulaire
3. Microscope à fluorescence
Utilisé pour détecter des molécules fluorescentes qui absorbent la lumière a une certaine longueur
d'onde et émettent à une longueur d'onde plus élevée. Des filtres vont permettre d'observer des
molécules luminescente sur fond noir. Il faut donc un microscope qui soit doté de 2 filtres:
• le premier qui ne laisse passer que la longueur d'onde d'absorption
• le deuxième qui ne laisse que la longueur d'onde d'émission.
La fluorescence peut-etre:
• Naturelle, émise par certains constituants de la cellule
• Rapportée par des molécules fluorescentes fixées, ou molécule utilisé comme detecteur:
Immunofluorescence.
On peut utiliser deux enzymes:
• Fluoriscine: qui absorbe le Bleu et émet dans le Vert.
• Rhodamine: absorbe Vert et donne du Rouge.
4. Microscope Confocale
Le microscope à fluorescence detecte la fluorescence de l'endroit sur lequel on a mis au point ainsi
que tout l'environnement l'entourant (comprendre: les différentes couches), il y aura alors un
bruit de fond.
Le microscope confocale ne recueille que la lumière émise au point sur
lequel on s'est focalisé. La lumière émise va traverser l'ouverture d'un
diaphragme (ouverture confocale) qui ne laissera traverser que la
lumière venant d'un point précis.
Le laser va balayer l'objet point par point sur un plan, puis par plan
successif. Un ordinateur va recréer l'image finale. En chaque point,
l'image sera nette.
L'épaisseur d'une couche est au minimum de 5 microns,
ainsi, certains détails peuvent échapper à l'observateur.
L'ensemble de ces systèmes ont une limite: la résolution qui est limité par:
• la longueur d'onde de la lumière
• du système optique
• de l'indice de réfraction du milieu entre la préparation et l'objectif (Note: les Objectifs à
Immersion sont baignés dans l'huile et permettent un agrandissement x100)
• le cône de lumière entre l'objet et la lentille (donné par la moitié de l'angle, soit ~70°)
La résolution maximale théorique est de 0,2 microns, or les organites ont une taille inférieure à 0,2
microns.
PCEM1 - Biologie Cellulaire 4/10 www.ProteinesPlus.frCours 01 - Les méthodes d'observation en Biologie Cellulaire
II. La microscopie Electronique (ME)
Elle est similaire au microscope optique, sauf qu'elle utilise des Electrons et les lentilles sont
électromagnétiques. La colonne surmontant l'installation est occupée par une cathode qui enverra
les electrons, un système de refroidissement, et un système permettant de faire le vide (car les
électrons ne voyagent pas dans l'air).
Le grossisement est reglé par les tensions appliquées sur les lentilles. Il y aura:
• Une lentille de condensation
• L'échantillon
• Une lentille d'objectif
• Le projectif qui projète l'image sur un écran fluorescent
• En dessous il y aura une plaque qui permet la capture d'image.
Les contraintes sont déterminées par:
• l'épaisseur de l'objet qui doit être encore plus fin que 5 microns
• les électrons font des contrastes et pas des colorations, donc l'image sera en noir et blanc.
1. Comment avoir une coupe ultrafine de 60-100 nm d'épaisseur ?
On peut observer les organites qui doivent être fixé très rapidement, sinon ils seront détruit.
➢ Fixation au Glutaraldéhyde, qui fixe les protéines très rapidement
➢ Post fixation à l'Acide Osmique qui fixe les lipides.
➢ Déshydratation en passant par un solvant
➢ Le bloc doit être très rigide pour pouvoir être coupé finement, il y a donc une inclusion en
résine: Epon ou Araldite semi-liquide à froid.
➢ La résine est chauffé à 60°C environ où elle polymérise
➢ L'échantillon enrobé sera de l'ordre du millimètre cube
➢ On découpe l'échantillon avec un Ultra-microtome plus sensible (avec un pas de 50nm).
Le couteau est fait de diamant et dont le fil fait 3mm.
➢ La coupe est maintenant ultra-fine avec une épaisseur de 100nm.
➢ Les coupes sont regroupées sur des grilles à maillage adapté.
La coupe flotte à la surface d'une cuve d'eau où elle sera récuperée
Avant de couper avec l'ultramicrotome, on vérifie que l'échantillon est interessant en faisant une
coupe semi-fine. La coupe semi-fine aura une épaisseur de 0.5 microns, et pourra être colorée et
observé au Microscope Optique.
2. Comment augmenter la visibilité ?
On peut augmenter le contraste en utilisant les sels de métaux:
• Citrate de Plomb
• Acetate d'Uranyle
Les électrons sont arretés par les métaux et l'image en tiendra compte
a. Coloration négative
Coloration de petits objets isolés et déposés sur une
grille recouverte d'un filtre.
Le produit de contraste vient se mouler autour des
objets, le MeT affichera alors une image en négatif
(en marquant les contours).
PCEM1 - Biologie Cellulaire 5/10 www.ProteinesPlus.frCours 01 - Les méthodes d'observation en Biologie Cellulaire
b. Ombrage métallique
Il met en évidence de petits éléments dans une cellule (ex: molécules protéique de la membrane
cellulaire). Le métal chauffé se dépose sur un coté de la structure.
Une fois déposé, il faudra solidariser la structure en appliquant un filtre transparent. L'échantillon
pourra ensuite être détruit, et on ne gardera que la réplique qu'on passera au MET. On aura alors
une image en contraste avec impression de relief.
L'ombrage métallique est souvent associé aux techniques suivante:
1. La cryofracture
Un échantillon est trempé dans de l'Azote liquide (pour le refroidir), puis cassé à l'aide
d'un couteau. La cassure se fera préférentiellement entre les deux feuillets d'une membrane.
Ainsi les faces internes des deux feuillets de la membrane pourront être examiné. Grâce à
l'ombrage métallique on pourra déterminer la concentration en protéines !
2. Le cryodécapage
On sublime l'eau du cytoplasme et on fait ainsi apparaître une partie du feuillet externe
de la membrane.
Cette technique est utilisé pour faire apparaître le Cytosquelette.
Le pouvoir de résolution du MET est dépendant de la longueur d'onde de l'electron.
Résolution maximale théorique : 0.1 nanomètre.
Grossissement: 3 000x (une partie de la cellule) à 100 000x (focalisation sur une structure précise,
dont les membranes).
PCEM1 - Biologie Cellulaire 6/10 www.ProteinesPlus.frCours 01 - Les méthodes d'observation en Biologie Cellulaire
III. La microscopie Electronique à Balayage(MEB)
Destiné à l'étude des surface cellulaire, il permet de voir la forme des objets. Son
principe de base est similaire au microscope à transmission.
Source d'electron: cathode, série de lentilles (voir disposition précedente)
Les cellules entières seront fixées puis métallisées (afin de maintenir la forme de la cellule), et le
tout sera balayé par un faisceau d'electron.
Lors du balayage, il y aura une réflection des électrons, qui seront detecté par un detecteur à
scintillation et transformation en impulsion electrique par un photomultiplicateur. Ces impulsions
electrique seront transformé.
L'image affiché sera en relief.
Ce MEB ne permet que de voir des surfaces et non la structure interne des cellules.
IV. Que peut-on observer
➢ Une observation générale
Coloration signalétique par Hematoxyline-eosine.
Etudes de cellules vivantes, utile pour les mitoses, avec le Microscope à contaste de
phase.
Les organites intracellulaires, le MET est requis.
Observer les différenciations des surfaces: le MEB.
➢ Localisation moléculaire
Histochimie: permet de mettre en évidence dans les cellules des constituant chimique
spécifique (Glucides, acides nucléiques, lipides....) la réaction histochimique la plus
connue est la PAS (Acide Periodique de Schiff).
Dans cette réaction, on oxyde les glycol par l'acide Periodique transformé alors en
Aldéhyde, puis on met en évidence les aldéhydes par le réactif de Schiff.
La coloration est Rouge Pourpre.
Histoenzymologie: permet de travailler en MO et ME. Detecte dans une cellule une
enzyme spécifique par la formation d'un produit à partir d'un substrat que l'on ajoute à
la préparation.
Au MO, le produit devra être coloré
ex: mise en evidence de la Peroxydase par le DAB (di amino benzidine)
Au ME: localisation de phosphatase acide dans les lysosomes, actif à un pH=5.
Précipité de phosphate de plomb visible au ME
Dans la plupart des cas il faut des fixations légères, donc tissus frais et qui a maintenu
l'activité enzymatique.
PCEM1 - Biologie Cellulaire 7/10 www.ProteinesPlus.frCours 01 - Les méthodes d'observation en Biologie Cellulaire
Immunocytochimie / Immunohistochimie: permet de detecter des
molécules/protéines/glycoprotéines in situ en utilisant la reconnaissance spécifique
entre un anticorp et un antigène.
Les anticorps (ou immunoglobuline) sont produit par les plasmocytes, qui sont le stade
de différenciation terminale des lymphocytes B.
Ces immunoglobulines sont de 5 variétés IgG IgA IgM IgE. Seul les IgG seront utilisées
Chaque Ig reconnaît un Epitope sur l'Ag ou site antigènique, formé par quelques acides
aminés. Une protéine aura donc de nombreux site antigènique.
Les anticorps peuvent être:
• Polyclonaux: ceux utilisés au départ, produit dans le cas d'une infection.
• On injecte à un animal (lapin) la protéine contre laquelle on veut produire des
anticorps. Cette protéine aura plusieurs sites antigèniques.
• L'animal développe plusieurs clones de LymphocytesB, chaque clone
reconnaissant un site antigènique différent.
• On prélève ensuite le sang.
• On cherche l'Ac spécifique par réaction: Ag-Ac.
• On prend la protéine et la fixe sur des billes placées dans une colonne de
purification
• On fait couler le sérum de l'animal sur ces billes
• Les Ac non spécifique tomberont, alors que les Ac spécifiques seront
retenus
• Il suffira de détacher les Ac pour avoir le produit purifié
C'est la manière la plus rapide
• Monoclonaux: Lorsqu'on utilise un Ac monoclonal, on aura une seule variété d'Ac,
provenant d'un seul clone.
• On injecte à l'animal (la souris) la protéine, l'animal développera des clones
dirigés contre les épitopes.
• On prélève les LymphocytesB (de la rate généralement) qu'on met en culture.
Cependant les lymphocytes ne se multiplient pas.
• Il faut les hybrider avec des cellules à capacité de multiplication infinie (dérivée
de tumeur/myelome). On obtient des lymphocytes qui vont être détruit, d'autre
seront hybrides, et d'autre uniquement tumorale, ces dernières devront être
eliminé via un milieu dans lequels ils ne pourront pas proliferer.
• Les cellules obtenues pourront créer des anticorps spécifique et proliferer.
• Il faut ensuite tester differents hybrides: on les repiquent individuellement et
on teste leur réactivité.
• On aura alors production de manière infinie du même anticorps.
On les utilise pour purifier des cellules (tri cellulaire), pour purifier des protéines (avec
le système de billes), expérimentation pour neutraliser les protéines in vivo, pour voir
l'effet des protéines sur certains mécanismes.
On les utilise aussi en thérapeutique, les Ac seront dirigé contre des recepteurs
membranaires (recepteurs aux facteurs de croissances) qui sont présent sur des
cellules cancereuses.
La detection in situ d'un Ag localisé dans une cellule est rendue possible grâce à la
réaction spécifique Ag-Ac. Il faut donc avoir une préparation (coupe en paraffine, coupe
au cryostat, coupes flottantes obtenues à partir d'un tissu fixé mais non enrobé par de
la paraffine).
Si on fait une congélation il y aura apparition de cristaux qui vont détruire la cellule, on
doit d'abord Cryoproteger en utilisant une solution élevée de Sucrose.
Le Vibratome permet de faire des coupes sans congélations, la lame de rasoir
vibrante avancant dans le tissu.
Les coupes seront receuillies dans un milieu qui permettra de faire la réaction
cytochimique.
PCEM1 - Biologie Cellulaire 8/10 www.ProteinesPlus.frCours 01 - Les méthodes d'observation en Biologie Cellulaire
On doit utiliser des marqueurs visibles: Fluorochrome, Enzyme (peroxydase), DAB
(coloré ET dense aux electrons), Or colloïdale dense aux électrons à diamètre variable
allant de 1-20 nm qu'on peut encore amplifier la réaction avec des sels d'argent.
La réaction pourra se faire de 2 manières:
• Méthode directe: L'Ac spécifique ou primaire, va directement être couplé avec le
marqueur. Signal visible que lorsqu'on a une quantité importante d'Ag dans la
cellule.
Cette méthode est surtout utilisé en diagnostic, si un patient a un dépôt d'Ac dans
un endroit donné, et on utilisera des Ac animal dirigé vers l'Ig humaine
• Méthode indirecte par amplification de signal.
• Utilisation d'Ac secondaire marqué, dirigé contre le premier Ac fixé à l'Ag.
• Utilisation du système Streptavidine – Biotine. La streptavidine est une
glycoproteine qui pourra fixer 4 fois une petite vitamine: la biotine.
• L'Ac primaire spécifique sera monté d'un Ac secondaire portant de la biotine qui
cherchera à s'attacher avec de la Streptavidine (marquée).
• Contrainte: l'Ag doit être préservé et il faut travailler à des concentrations
adéquat de telle sorte que l'on ait un rapport du signal spécifique sur le bruit de
fond (réaction aspécifique) faible. Il faut aussi éviter des faux positif:
reconnaissance de molécules non-interessante.
Detections des acides nucléiques: Hybridation in situ, permettant de détecter des
séquences d'ADN ou d'ARN messager par la complémentarité des acides nucléiques.
Autoradiographie: utilisant des radio-isotopes (Tricium par ex). Ces particules vont
impressionner un film photographique placé au dessus de l'échantillon qu'on va
observer. L'échantillon devra être fait dans le noir (vu le developpement photo). A
chaque endroit où se trouve le radio-isotope, il y aura une image latente.
Cette technique est utilisé pour étudier la liaison entre une molécule et son recepteur:
on injecte la protéine/molécule marquée avec du radio-isotope, puis on attend que la
liaison se fasse entre la protéines et son recepteur, puis on prélève, on coupe et on
observe où s'est fixé la protéine, qui révèlera la position des recepteurs. Très utilisé
dans l'hybridation in situ et dans l'étude de la prolifération cellulaire.
Etude de la prolifération cellulaire: on injecte à un animal de la Thymidine Triciée
qui va s'incorporer dans l'ADN (dans les phases de synthèse de l'ADN et donc toute les
cellules en phase de synthèse vont l'incorporer). On relèvera alors le nombre de noyau
marqué par la Thymidine Triciée.
PCEM1 - Biologie Cellulaire 9/10 www.ProteinesPlus.frCours 01 - Les méthodes d'observation en Biologie Cellulaire
➢ Techniques de séparation et de purification concernant les cellules entière ou alors,
les organites cellulaire:
Le tri cellulaire: On cherche à obtenir une variété de cellules aussi pure que possible
pour l'étudier.
• On réduit d'abord le tissus en une suspension de cellules, grâce à des enzymes qui
élimine la MeC
• Séparer les cellules si elles sont jointives grâce à des Chélateurs du Calcium, car
celui çi intervient de manière importante dans les liaisons cellulaires.
• Les cellules sont de types variées. Parmis ces cellules on en cherche une seule
variété, il faut purifier cette préparation en utilisant des Ac qui reconnaitront de
manière spécifique des protéines portées uniquement par les cellules qui nous
interesse.
• Fixation des Ac sur un support et de passage de la préparation sur ce
support. Les cellules seront arreté sur les Ac, alors que les cellules en
suspension ne nous interesse pas.
• Cytomètre à flux (Cytomètrie de flux): les Ac portent cette fois çi un
fluorochrome, et seront couplé à leur Ag. La suspension sera passée dans une
colonne qui débouche sur un interstice d'un diamètre d'une cellule.
Les cellules passeront l'une après l'autre devant un detecteur de fluorescence
et l'appareil va pouvoir placer les cellules dans une goutelette chargé
positivement ou négativement en fonction de la detection du fluorochrome. Les
cellules passeront devant un deflecteur qui déviera les goutelettes chargée
dans un sens ou dans l'autre.
Système extrèmement rapide.
➢ Observation des organites:
Il faut fractionner les structures intracellulaires:
• Destruction des membranes cytoplasmique par broyage grâce à un piston qui
laminera le long de la paroi, on travaille généralement dans la glace pour préserver
les petites structures
• Eclatement des cellules dans un milieu hypotonique: Appel d'eau dans la cellule
qui amène à l'éclatement, c'est un choc Osomotique
• Utilisation d'Ultrason
L'homogénat obtenu devra maintenant être fractionné par Ultra centrifugation dite
Différentielle ou on augmente la vitesse et le temps.








 La plupart des produits dits dietsdoivent leur goût sucré à l'
La plupart des produits dits dietsdoivent leur goût sucré à l'